Il représente le sushi japonais : le thon. Cette espèce n’est pas consommée seulement en sushi, mais aussi sous forme de conserve. Le thon est une ressource halieutique très importante. Son énorme corps (jusqu’à 2m) et ses mystères attirent les chercheurs. D’autre part, la gestion du stock de thon est devenue un sujet important au sein de coopération internationale à cause de la baisse remarquable de ses captures. Notre laboratoire effectue ses recherches sur la migration du thon par des marquages d’individus, surtout au niveau des espèces : thon blanc, thon rouge du Pacifique et le thon à nageoire jaune. Afin d’étudier les effets des fluctuations de leur environnement sur la ressource de thon, des analyses chimiques des œufs, larves et otolithes sont utilisés.
1. Pacific Bluefin Tuna (Thunnus orientalis) - Migration Ecology/ Le thon rouge du Pacifique (Thunnus orientalis)-Ecologie de sa migration
Le thon rouge du Pacifique est l’une des espèces cible commerciale principale pour la qualité de sa chaire (Image 1). Sa capture atteint 10 000 tonnes annuelles au Japon. Cependant, ses captures sont en baisse depuis les années 70. En tant que premier marché du thon rouge, une gestion correcte de sa ressource est un sujet très important pour nous. Leur migration à grande échelle nécessite un plan de gestion internationale et une connaissance approfondie de leur mécanisme migratoire. Nous utilisons des marquages (Image 2) pour l’étude de leur comportement. Notre but est de trouver comment le thon rouge s’adapte à des eaux tempérées en mesurant le comportement migratoire des jeunes thons dans le Pacifique Nord. Ces mesurent incluent les conditions environnementales et physiologiques des échantillons, ainsi que les facteurs physiques et biologiques influençant leur comportement migratoire.
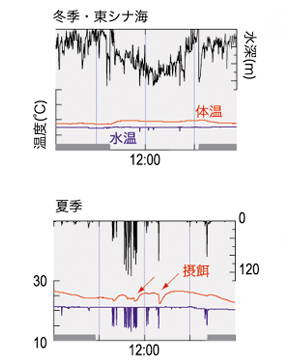
Nos recherches ont révélé que le thon rouge du Pacifique dans la mer de Chine a un cycle circadien lors de sa migration verticale : il reste dans les couches superficielles, près de la surface, pendant la nuits en hivers et descend vers des eaux plus profondes pendant le jour (Figure 1). Certains individus migrent au large de la région Nansei Shoto, là où une stratification des températures a été observée. Leur cycle circadien y est particulièrement marqué ; cette stratification semble donc être un élément clé au cycle. D’autre part, des individus se déplaçant au larges du Sanriku restent dans des zones chaudes (Image 3). Dans cette région, la stratification est assez faible et les thons ont tendance à rester au niveau des couches superficielles et descendre moins en profondeur que les individus de la mer de Chine. De plus, leur fréquence d’alimentation a été calculée en détéctant les chutes de température corporelle, suivant l’ingestion d’aliments. Ces analyses indiquent que les individus au large de Sanriku s’alimentent plus souvent par jour que rapport au individus de la mer de Chine. Ces résultats révèlent que cette espèce change de stratégie alimentaire en fonction de sa localisation : recherche de nourriture en-dessous la thermocline de la mer de Chine ou recherche horizontale au large de Sanriku (Figure 2-1 & 2-2). Les archives de marquages ont apporté des données détaillées sur la compréhension de leur écologie. Les modifications spaciales et saisonnières de la température de l’eau et les stratifications influencent fortement la migration verticale du thon rouge du Pacifique.
 |
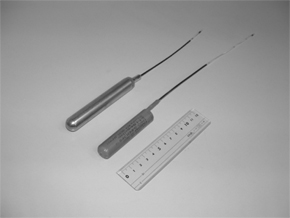 |
| Image 1-Migration du thon rouge du Pacifique | Image 2-Marqueurs |
 |
 |
| Figure 1-Profondeur de nage des thons rouges du Pacifique dans la mer de Chine | Image 3-Image satellite des thons rouges dans un tourbillon chaud au large de Sanriku |
 |
 |
| Figure 2-Recherche de nourriture au large de Sanriku | Figure 3-Recherche de nourriture dans la mer de Chine |
2. Les mécanismes de thermorégulation du thon
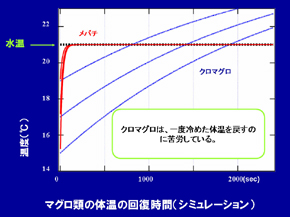
Le thon rouge du Pacifique a une température corporelle plus élevée que celle de l’eau dans laquelle il se trouve. De plus, la différence entre ces températures tend à s’agrandir lorsque la température de l’eau chute. Des modèles mathématiques indiquent qu’une inertie thermique et une forte production de chaleur contribue à maintenir cette température corporelle élevée. Des calculs ont révélé que les thons rouges du Pacifique atteignent des productions de chaleur interne aussi élevées que les mammifères. En effet, ils peuvent maintenir leur température péritonéale lorsqu’ils évoluent dans un environnement froid. Cependant, leur capacité à récupérer des températures corporelles basses grace à la température de l’eau dans laquelle ils se trouvent n’est pas bonne (Image 3). Pour cette raison, ils restent dans les eaux chaudes et entrent dans les eaux froides seulement pour chercher de la nourriture. Le thon rouge du Pacifique gère sa thermorégulation via des modifications comportementales. Le taux d’insolation a aussi un effet sur leur migration verticale : la fréquence de migration verticale diminue lorsque l’insolation diminue.
Plus le thon grandit, plus son corps atteint des températures élevées par rapport à l’eau que l’entoure. Cependant, la température moyenne de son corps ne dépasse pas 30°C. Des modèles mathématiques indiquent que sa vitesse de production de chaleur et son taux de transfert de chaleur diminuent lorsqu’il grandit. La température corporelle ne doit pas atteindre la valeur létale. Grâce à ces capacités, il peut rester dans des eaux tempérées et grandir jusqu’à obtenir une taille importante faisant de lui l’un des plus grands poissons du monde.
Nous étudions aussi maintenant le thon à nageoire jaune(T. albacares)qui est une grande espèce vivant dans des eaux tropicales. Nous l’étudions avec les mêmes méthodes (marquages) pour nous éclairer sur ses mécanismes migratoires (Image 4).
 |
 |
| Figure 4-Temps de récupération du thon rouge | Image 4-Marquage d'un thon à nageoires jaunes |
3. Influence des fluctuations de l’environnement marin sur les thons
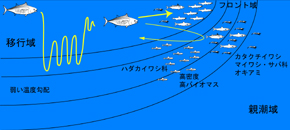
Les thons rouges du Pacifique ont une aire de fraye particulière comparée à leur large répartition dans l’océan Pacifique : au Sud de l’océan, en-dessous de 30° Nord et à l’Est de Kuroshio. La saison de fraye a lieu de mai à juin autour des îles Nansei, de juin à juillet au large et Sud de l’île principale du Japon, et de juillet à août dans la mer du Japon. Ils grandissent dans les couches superficielles de la mer à des températures entre 25 et 26°C. La thermocline est de 50m ou moins profond. Par rapport à leur distribution, l’aire de fraye est très limitée. Nous recherchons comment les fluctuations de l’environnement marin affectent le comportement et la reproduction des thons (Images 5).
Quelle est l’influence des variations de temperatures sur les oeufs et les larves de thons rouges du Pacifique? Des élevages expérimentaux ont montré que leur survie et croissance seraient affectées négativement lorsque l’eau atteindrait des températures trop hautes ou trop basses. Le rapport RNA/DNA, la consommation d’oxygène et le taux de déformation des larves indiquent aussi une influence des augmentations de la température de a surface de l’eau.
De plus, les otolithes des thons rouges du Pacifique sont aujourd’hui considérés comme des indicateurs des fluctuations de l’environnement marin. Le taux d’isotope stable d’oxygène des otolithes est comme un enregistrement des modifications de température et environnementales. Si les différences environnementales des aires de fraye sont enregistrées dans les otolithes, la structure de la population de thons rouges du Pacifique peut être analysée en détail. Nos recherches peuvent apparaitre comme un tournant décisif dans la gestion du thon rouge du Pacifique.
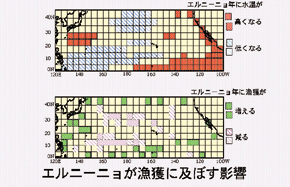
Les thons blancs (T. alalunga)du Nord du Pacifique ont été ciblés pour des analyses des facteurs environnementaux sur l’écologie de la migration du thon, sa distribution dans le temps et l’espace depuis sa reproduction jusqu’à sa maturité. Les résultats montrent que leurs routes de migrations changent lorsqu’ils atteignent l’âge de 6 ans, formant un cycle tournant dans un sens antihoraire. Cependant certains individus plus âgés du centre du Pacifique se déplacent vers l’Est du Pacifique lorsqu’a lieu El Niño (Figure 4).
 |
 |
| Image 6-Larves de thons rouges du Pacifique | Image 5-Effets d'El Niño sur les thons blancs |
References
- Kimura, S., Nakai, M. and Sugimoto, T. (1997) Migration of albacore (Thunnus alalunga) in the North Pacific Ocean in relation to large oceanic phenomena. Fisheries Oceanography, 6, 51-57.
- Kitagawa, T., Sartimbul, A., Nakata, H., Kimura, S. and Yamada, H. (2006) Effect of water temperature on habitat use of young Pacific bluefin tuna, Thunnus orientalis, in the East China Sea. Fisheries Science, 72, 1166-1176.
- Kitagawa, T., Kimura, S., Nakata, H. and Yamada, H. (2006) Thermal adaptation of Pacific bluefin tuna Thunnus orientalis to temperate waters. Fisheries Science, 72, 149-156.
- 北川貴士.(2006) バイオロギングによるクロマグロの行動生態研究の現状.テレメトリー-水生動物の行動と漁具の運動解析-.山本勝太郎,山根 猛,光永 靖編,恒星社厚生閣,45-55.
- 北川貴士.(2005) クロマグロの遊泳行動とそれに及ぼす物理環境要因.月刊海洋,6:海洋動物の認知システムと回遊メカニズム,416-421.
- 北川貴士.(2005) マグロ類の遊泳と回遊.海の生物資源-生命は海でどう変動しているか-,海洋生命系のダイナミクス・シリーズ第4巻,渡邊良朗編,東海大学出版,37-53.
- 北川貴士.(2005) クロマグロ体温と環境水温の熱交換モデル.水産工学,41,257-260.
- Kitagawa, T., Kimura, S., Nakata, H. and Yamada, H. (2004) Diving behavior of immature Pacific bluefin tuna (Thunnus thynnus orientalis) for feeding in relation to seasons and areas: the East China Sea and the Kuroshio-Oyashio transition region. Fisheries Oceanography, 13, 161-180.
- Kitagawa, T., Kimura, S., Nakata, H., Yamada, H. (2004) Overview of the research on tuna thermo-physiology using electric tags. Memoirs of the National Institute of Polar Research, Special, Issue 58, 69-79.
- Kitagawa, T., Kimura, S., Nakata, H. and Yamada, H. (2004) Effect of light intensity on the vertical movement of Pacific bluefin tuna Thunnus orientalis. Zoological Science, 21, 1258-1259.
- 北川貴士.(2004) クロマグロの遊泳行動とそれに及ぼす海洋要因.海流と生物資源,杉本隆成編著,成山堂,224-236.
- Kitagawa, T., Kimura, S., Nakata, H. and Yamada, H. (2003) Diving patterns and performance of Pacific bluefin tuna (Thunnus thynnus orientalis) as recorded by archival tags. Otuchi Marine Science, 28, 52-58.
- Kitagawa, T., Kimura, S., Nakata, H. and Yamada, H. (2003) Adaptation mechanisms of Pacific bluefin tuna to temperate waters. Zoological Science, 20, p1526.
- 北川貴士.(2003) クロマグロの日周鉛直行動とその意味ミ東シナ海で再捕されたクロマグロ未成魚の遊泳状況. アクアネット,6,(6),31-35.
- Kitagawa, T., Nakata, H., Kimura, S., Sugimoto, T. and Yamada, H. (2002) A review: Analysis of tunas behavior using of acoustic, archival and pop-up tags methods. Otsuchi Marine Science, 27, 1-6.
- Kitagawa, T., Nakata, H., Kimura, S. and Yamada, H. (2002) Diving behavior of immature Pacific bluefin tuna (Thunnus thynnus orientalis) recorded by an archival tag. Fisheries Science, 68, Supplement I, 427-428.
- Kitagawa, T., Nakata, H., Kimura, S., Sugimoto, T., Yamada, H. (2002) Differences in vertical distribution and movement of Pacific bluefin tuna (Thunnus thynnus orientalis) among areas: the East China Sea, the Sea of Japan and the western North Pacific. Marine and Freshwater Research, 53, 245-252.
- 北川貴士.(2002) クロマグロ仔稚・未成魚の温帯水域への輸送・回遊. 月刊海洋,号外31,黒潮の生物輸送と生産機能,152-157.
- Kitagawa, T., Nakata, H., Kimura, S. and Tsuji, S. (2001) Thermoconservation mechanism inferred from peritoneal cavity temperature recorded in free swimming Pacific bluefin tuna (Thunnus thynnus orientalis). Marine Ecology Progress Series, 220, 253-263.
- Kitagawa, T., Nakata, H., Kimura, S., Itoh, T., Tsuji, S. and Nitta, A. (2000) Effect of ambient temperature on the vertical distribution and movement of Pacific bluefin tuna (Thunnus thynnus orientalis). Marine Ecology Progress Series, 206, 251-260.
- 溝呂木奈緒.(2009) 地球温暖化に伴う水温上昇がクロマグロ(Thunnus orientalis)の初期生残に及ぼす影響.東京大学大学院新領域創成科学研究科自然環境学専攻,修士論文.
- 中井宗紀・木村伸吾・杉本隆成.(1994) ビンナガの回遊.月刊海洋,26,547-551.